
Chloridoideae. -- The subtribe Chloridoideae, consisting of 146genera and 1626 species (Table 2) from dry regions of the tropics, has been the subject of a number of phenetic and phylogenetic studies involving both the whole tribe as well as sections of it. Early work was phenetic, a good example being a study of the Eragrostideae (Phillips 1982) based mainly on inflorescence and spikelet characters. This was followed by the research that combined phenetic and cladistic methods (van den Borre & Watson (1994) for Eragrostis and van den Borre & Watson (1997, 2000) for the whole subfamily based on morphological and anatomical characters). A cladistic study based on morphological characters was published for 38 New World genera of the Eragrostideae (Peterson, 2000) and more recently cladistic ork has been based on molecular data (Hilu & Alice, 2001; Columbus et al. 2003; Columbus et al., 2007; Liu et al. 2007; Peterson et al. 2007; Peterson et al. 2010; Peterson et al. 2012).
The subfamily, while having affinities with the Arundinoideae, has been presented as being monophyletic on both morphological (Kellogg & Campbell, 1987) and molecular (Hilu & Alice, 2001) grounds. The only synapomorphic morphological character has been stated as the possession of “chloridoid” bicellular microhairs with an inflated apical cell (Tateoka et al., 1959), although there appears a difference of opinion of whether these chloridoid microhairs also occur in non-chloridoid groups (Kellogg & Campbell, 1987; Jacobs, 1987). The “panicoid” type of microhairs can also occur in the Chloridoideae, although they are not as common as the “chloridoid” type.
An early published phylogenetic study of the whole subfamily (Hilu & Alice, 2001) sampled 74 species belonging to 56 genera and based on the plastid gene matK. The resultant consensus of 5,800 equally parsimonious trees show a topology of four main clades which do not align at all well with some of the traditional subtribes in this subfamily. Some of the genera are shown to be paraphyletic or polyphyletic; some morphological feature are shown to be synapomorphic for some clades within the topology, but many more are homoplasious. In a classification of 72 genera of New World chloridoids (Peterson et al., 2007), a new chloridoid classification was proposed, based on the synthesis of previous morphological and molecular work, with the recognition of three tribes, Cynodonteae, Eragrostideae and Zoysieae. Of interest in this classification is the separation of Eragrostis in the Eragrostideae and Sporobolus in the Zoysieae, whereas a previous molecular study based on ITS had included some species of Eragrostis in a Sporobolus clade (Ortiz-Diaz & Culham, 2000). Chloridoid genera of particular interest in terms of their recognition in the chloridoids include Thellungia Stapf and Stiburus Stapf. Both were placed in Eragrostis on the basis of a phenetic morphological study (Phillips, 1982) but on the basis of molecular studies both are shown to maintain a separate and unique identity. Thellungia was shown to have a closer relationship with Sporobolus than with Eragrostis, based on molecular work (Ortiz-Diaz & Culham, 2000, Hilu & Alice, 2001); Stiburus is recognized it its own right and related to Neeragrostis (Bell & Columbus, 2008), rather than aligning with an Eragrostis clade. However the genera Eragrostis and Sporobolus, two of the largest in the Chloridoideae, have both been shown not to be monophyletic on the basis of molecular work; Eragrostis is polyphyletic (van den Borre 1994, Bell & Columbus, 2008) and Sporobolus is paraphyletic (Ortiz-Diaz & Culham 2000, Bell & Columbus 2008).
The chloridoid subtribe Boutelouinae, mainly occurring in the New World, was until fairly recently, presented as consisting of 16 genera, all fairly narrowly circumscribed (Clayton & Renvoize 1986). The core genus Bouteluoa Lag. was revised traditionally by Gould (1979), with two subgenera, subgenus Bouteluoa and subgenus Chondrosium; he had earlier presented a case for keeping them as separate genera (Gould 1963), a position followed by Clayton and Renvoize. Allied to Bouteloua, which has bisexual inflorescences, are a few dioecious genera (Buchlominus Reeder, Reeder & Rzedowski, Cyclostachya J. & C. Reeder) or genera that have species which are monoecious or dioecious (Pringlechloa Scribner, Opizia J. & C. Presl, Buchloe Engelm., Soderstromia C.V.Morton). In recent times however most of these narrow defined allied genera have been placed in a widely circumscribed Bouteloua following phylogenetic analysis based on two molecular sequences of both nuclear and chloroplast DNA (ITS and trnL-F) (Columbus et al., 1998; Columbus, 1999; Columbus, et al. 2000).
The endemic chloridoid Australian tribe Triodieae (the Spinifex grasses), was originally classified (Jacobs, 1971) into two large genera, Triodia R.Br. and Plectrachne Henrard, on the basis of whether the lemmas were awned or not. In 1985 two more genera, Symplectrodia Lazarides and Monodia S.W.L. Jacobs, were described (Lazarides, 1985; Jacobs, 1985) based on morphological characters. Symplectrodia differs from the other genera by three characters and Monodia differs by two. A recent revision (Lazarides, 1997) does not uphold the Triodia - Plectrachne division, on the basis that the traditional spikelet characters formerly used to distinguish the genera break down. Furthermore leaf anatomical characters cut across the traditional Triodia - Plectrachne classification (Burbidge, 1946), with the recognition of “soft” and “hard” spinifex types depending on whether stomatal grooves are located on one or both leaf surfaces. A molecular analysis in the Trioidieae, based on the ITS region, was undertaken on 27 representative taxa and three outgroups (Mant et al., 2000). Representing the tribe were 17 species of Triodia s.s., 7 species of Triodia formerly placed in Plectrachne, 2 species of Symplectrodia and the monospecific Monodia. The strict consensus of 22 most parsimonious trees shows strong support for monophyly of the Triodieae, but within the ingroup anatomical characters give greater support to the groupings than do those of spikelet morphology. The topology shows 6 clades, of which one includes species with a “soft” leaf anatomy and another a clade of four southern temperate species of Triodia. Both include members of species formerly placed in Triodia and Plectrachne. Monodia is shown to be apomorphic and Symplectrodia is in a clade that makes two other species of Triodia paraphyletic. Complete sampling across the whole range of the tribe is necessary before any new classification is proposed.
The most recent classification of the Chloridoideae is that of Peterson, Romaschenko & Johnson (2010), who produced a comprehensive molecular phylogeny for the subfamily that include 254 species in 99 genera. These are delimited into four tribes: Triraphideae, Eragrostideae, Zoysieae, and Cynodonteae; in addition there are 26 genera that have not been sampled moleculularly and that are present placed as Chloridoideae incertae sedis Subsequent to the latter publication is the description of a new tribe Centropodieae basal to all other chloridoids (Peterson, Romaschenko, Barker & Linder 2011) that includes the genera Centropodia and a new genus Ellisochloa. Traditional morphological characters in this subfamily are proving to be evolutionary labile, with some major genera (Leptochloa, Chloris and Sporobolus) being polyphyletic. Eragrostis appears to be monophyletic when genera such as Ectrosia and Pogonarthria are subsumed within it. When the polyphyletic genus Leptochloa was studied using five plastid and one nuclear marker (Peterson et al. 2012), it was reclassified into 5 separate genera on the basis of the new lineages discovered. One of the new genera (Trigonochloa) has been revised (Snow & Peterson 2012) and others are to follow (Snow pers. comm.).
.

Paul M. Peterson, Konstantin Romaschenko & Gabriel Johnson (2010). Molecular Phylogenetics and Evolution 55, Fig.3.

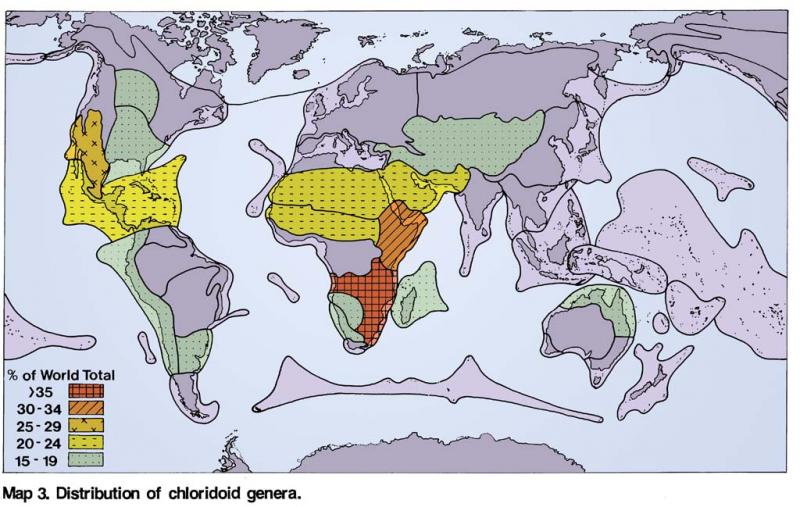
Distribution of Chloridoideae (Hartley 1960)
Simon & Jacobs 1990
Ce Centropodieae CIS Chloridoideae incertae sedis Cy Cynodonteae Er Eragrostideae Tr Triraphideae Zo Zoysieae
Cy Acrachne
Cy Aeluropus
CIS Afrotrichloris
Er Allolepis
Cy Apochiton
Cy Astrebla
Cy Austrochloris
Cy Bewsia
Cy Blepharidachne
Cy Bouteloua
Cy Brachyachne
Cy Brachychloa
Zo Calamovilfa
Cy Catalepis
Ce Centropodia
Chaboissaea
Cy Chloris
Cy Chrysochloa
Er Cladoraphis
Cy Cleistogenes
Cy Coelachyrum
Er Cottea
Cy Craspedorhachis
Zo Crypsis
Cy Ctenium
Cy xCynochloris
Cy Cynodon
Cy Dactyloctenium
CIS Daknopholis
Er Dasyochloa
CIS Decaryella
CIS Desmostachya
Cy Dignathia
Cy Dinebra
Cy Distichlis
CIS Drake-Brockmania
Er Ectrosia
Cy Eleusine
Ce Ellisochloa
Er Enneapogon
Cy Enteropogon
Er Entoplocamia
Cy Eragrostiella
Er Eragrostis
Cy Erioneuron
Cy Eustachys
CIS Farrago
Er Fingerhuthia
Cy Gouinia
Cy Gymnopogon
CIS Habrochloa
CIS Halopyrum
Er Harpachne
Cy Harpochloa
Cy Heterachne
CIS Heterocarpha
Cy Hilaria
CIS Hubbardochloa
CIS Indopoa
Cy Jouvea
CIC Kampochloa
Er Kaokochloa
CIS Leptocarydion
Cy Leptochloa
Cy Leptothrium
Cy Lepturidium
CIS Lepturopetium
Cy Lepturus
Cy Lintonia
CIS Lophacme
Cy Lopholepis
Cy Melanocenchris
Cy Microchloa
Cy Monelytrum
Er Monodia
Cy Mosdenia
Cy Muhlenbergia
Cy Munroa
CIS Myriostachya
Cy Neesiochloa
Cy Neobouteloua
Cy Neostapfia
CIS Neostapfiella
Tr Neyraudia
CIS Ochthochloa
Cy Odyssea
Cy Orcuttia
Cy Orinus
Cy Oropetium
Cy Oxychloris
Cy Pappophorum
Cy Perotis
Er Pogonarthria
CIS Pogonochloa
Zo Pogononeura
Cy Polevansia
CIS Pommereulla
Er Psammagrostis
CIS Pseudozoysia
CIS Psilolemma
Cy Rheochloa
Er Richardsiella
Cy Saugetia
Cy Schedonnardus
Er Schmidtia
Cy Schoenefeldia
Cy Sclerodactylon
Cy Scleropogon
CIC Silentvalleya
Cy Sohnsia
Zo Spartina
Zo Sporobolus
Cy Steirachne
Cy Stiburus
Cy Swallenia
Cy Symplectrodia
CIC Tetrachaete
Cy Tetrachne
Cy Tetrapogon
Zo Thellungia
Cy Trikeraia
Cy Toliara
Cy Tragus
Cy Trichloris
Cy Trichoneura
Cy Tridens
Cy Triodia
Cy Triplasis
Cy Tripogon
Tr Triraphis
Cy Tuctoria
Er Uniola
Zo Urochondra
Cy Vaseyochloa
Er Vietnamochloa
CIS Viguierella
Cy Willkommia
Zo Zoysia